
'GAZE'
EVOLUZIONE GENETICA DI RETI NEURALI ARTIFICIALI PER IL CONTROLLO MOTORIO DI UN MODELLO BIDIMENSIONALE
OCCHI -TESTA
Dopo che ALIFE ha mostrato l’emergenza ed evoluzione di sensori, effettori e coordinazione sensori-motoria generale, GAZE mostra l’evoluzione di una coordinazione sensori-motoria specifica: quella occhi-testa, per seguire con lo sguardo un bersaglio mobile.
Qui l’obiettivo è mettere alla prova l’ipotesi che analoghe richieste ambientali (compiti influenzanti la fitness), ed analoghi equipaggiamenti e vincoli fisici, portano all’evoluzione di analoghi comportamenti e meccanismi neurali di controllo. L’indagine è quindi focalizzata a livello del controllo neurale, mentre i sensori (occhi e orecchie) e gli effettori (muscoli per muovere la testa e gli occhi) sono assunti come già presenti, già evoluti, e sono simulati a un livello più realistico rispetto ad ALIFE.
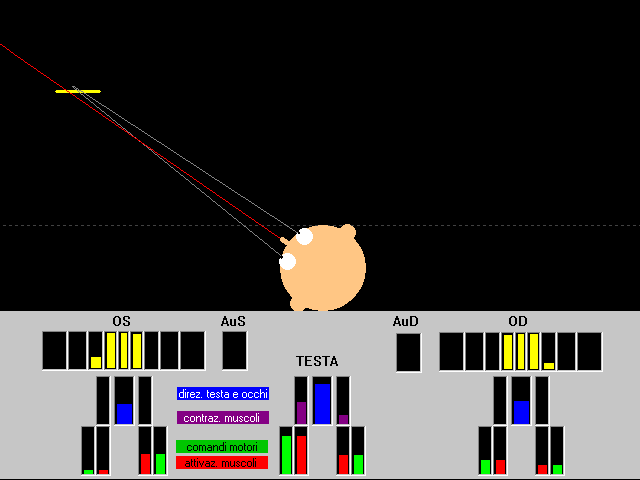
La simulazione assume un modello semplificato occhi-testa bidimensionale: una coppia di muscoli antagonisti permette la rotazione della testa sul piano orizzontale, due altre coppie di muscoli antagonisti simulano l’azione dei muscoli retti mediali e laterali oculari così che ogni occhio può ruotare indipendentemente orizzontalmente entro l’orbita. Due “retine” ricevono l’immagine di un oggetto lineare (bersaglio) che si muove casualmente davanti agli occhi in posizioni diverse con velocità variabile, e a volte emette un suono (simulato) che viene ricevuto dalle “orecchie”.
La rete neurale riceve input “visivi” dalle due retine, input “acustici” dalle due orecchie, informazioni “propriocettive” sugli angoli degli occhi e della testa e sulla tensione dei muscoli degli occhi e della testa; e genera output analogici di attivazione per quei medesimi muscoli.
Sensori
Le retine sono composte ciascuna di 9 campi recettivi (una “fovea” centrale e 4 recettori laterali simmetrici) parzialmente sovrapposti, ognuno dei quali informa della sua attivazione una unità di input della rete neurale.
Le orecchie ricevono brevi suoni emessi a intervalli casuali dal bersaglio, suoni che arrivano con intensità inversamente proporzionale alla distanza bersaglio-orecchio (le differenze di fase non sono prese in considerazione in questa simulazione semplificata), e ogni orecchio invia le sue informazioni ad una unità di input della rete neurale.
Il corredo sensoriale è completato da informazioni propriocettive dai muscoli e legamenti del collo e degli occhi, per un totale di 29 unità di input della rete neurale: 18 visive, 2 uditive, e 9 propriocettive.
Effettori
La testa e gli occhi sono fatti ruotare da due muscoli ciascuno, uno che fa ruotare in senso orario (verso destra) e l’altro in senso antiorario (verso sinistra). I due muscoli della testa simulano i muscoli sternocleidomastoidei, i due muscoli di ciascun occhio simulano i muscoli retti laterale e mediale. I punti di inserzione muscolare, le lunghezze dei muscoli, le rotazioni massime di testa e occhi, e il rapporto di massa occhi-testa, sono mantenuti conformi alla realtà biologica, così come pure la curva tensione-lunghezza dei muscoli, simulati con un modello a molla.
Ogni muscolo è azionato da una unità di output della rete neurale di controllo, la cui attivazione determina lo stato di tensione del muscolo corrispondente.
Rete neurale
La rete neurale è quindi costituita da 29 unità di input e 6 di output, fisse in quanto legate al sistema dei sensori ed effettori che in questo esperimento non è soggetto a mutamenti evolutivi; e da unità nascoste inizialmente in numero casuale (da 8 a 32 nei diversi individui), con connessioni casuali fra esse e con le unità di input e output, successivamente riconfigurate in numero e connessioni esclusivamente dalla selezione evolutiva (è lasciata all’evoluzione la selezione non solo dei pesi delle connessioni, ma anche dell’architettura delle reti).
Genotipo, fenotipo, algoritmo genetico
La codifica del fenotipo nel genotipo è attuata con una soluzione originale consistente nel suddividere il genotipo in due cromosomi separati, che consente di evitare l’uso – altrimenti obbligatorio con genomi a lunghezza variabile – di chiavi (etichette) per identificare le varie unità, e di marcatori per identificare i punti iniziale e finale di ogni segmento di genoma che definisce una unità: chiavi e marcatori che appesantiscono il genoma e la computazione, e non sono molto plausibili biologicamente.
Diversamente da ALIFE, in questo esperimento la selezione genetica e le interazioni tra gli individui sono simulate a un livello piuttosto astratto. La popolazione è costituita da un numero fisso e costante di 100 individui. In ogni generazione ciascun individuo è fatto vivere per 10000 turni di simulazione in modo indipendente dagli altri individui, e sono misurate le sue prestazioni nel seguire con lo sguardo il bersaglio lungo un percorso casuale identico per ciascun individuo di quella generazione. Alla fine della generazione le 100 prestazioni “in solitario” sono messe a confronto, e agli individui migliori è dato un numero di discendenti maggiore rispetto agli individui peggiori. Perciò, diversamente da ALIFE, le generazioni non si sovrappongono: i figli non vivono contemporaneamente ai genitori, ma sono generati alla fine della vita di quest’ultimi, in numero probabilisticamente proporzionale alla fitness del genitore.
Risultati
In 3500 generazioni (circa 90 ore su un comune PC) la popolazione ha raggiunto un errore medio nel seguire il bersaglio (differenza angolare tra la direzione della testa e degli occhi e il centro del bersaglio, media delle 100 reti in ogni generazione) di circa 5°.
Tale risultato è ottenuto tramite reti con 23 unità nascoste, valore consolidatosi fin dalla 500a generazione (con eccezione per singoli mutanti con 4 - 40 unità nascoste che non si sono mai affermati).
Più ancora dell’andamento dell’errore complessivo è interessante esaminare le caratteristiche del controllo sensori-motorio ottenuto. Per fare questo abbiamo analizzato in dettaglio la prestazione dell’individuo migliore alla generazione 4000.
Un primo test è consistito nel far seguire con lo sguardo un bersaglio che si muoveva lungo una traiettoria prestabilita con movimenti rapidi e lenti e con pause.
La rete si è mostrata capace di attuare efficacemente il controllo motorio desiderato, generando movimenti oculari rapidi coniugati (foveazione binoculare) e non coniugati (convergenza), e movimenti rapidi (saccadi) e lenti (inseguimento) corrispondenti alla posizione e velocità del bersaglio. Questo tipo di comportamento è risultato molto simile a quello di sistemi biologici (uomo e primati).
In particolare abbiamo osservato che, di fronte amovimenti rapidi del bersaglio, gli occhi si muovono velocemente così che lo sguardo raggiunge rapidamente la posizione approssimativa del bersaglio; poco dopo la testa inizia a muoversi e gli occhi compiono un movimento compensatorio retrogrado, armonico con la rotazione della testa, che li riporta in posizione primaria pur mantenendo lo sguardo fisso sul bersaglio. I tracciati di questi movimenti sono molto simili a quelli registrati da diversi autori in esperimenti nell’uomo e nei primati (Bizzi, E.: The Coordination of Eye-Head Movements. Sci Am. 231 (1974) 100-106) .
Se il movimento del bersaglio è abbastanza lento da poter essere seguito dalla testa non si osserva alcun movimento degli occhi, ancora similmente a quanto accade nei sistemi biologici.
Un secondo test è consistito nel ripetere il medesimo compito con la testa bloccata. La rete ha compensato efficacemente tramite ampi movimenti oculari, proprio come fanno i sistemi biologici. Questo risultato è particolarmente interessante in quanto va notato che la condizione “blocco della testa” non era mai stata sperimentata dalla popolazione in evoluzione.
Un terzo test analogo al precedente è stato effettuato bloccando gli occhi anzichè la testa. Anche in questo caso si è manifestato un comportamento compensatorio, almeno per movimenti del bersaglio abbastanza lenti da consentire l’inseguimento con la testa (di massa e inerzia ovviamente maggiore rispetto agli occhi).
Infine un quarto test è consistito nel variare le dimensioni del bersaglio. Mentre nei precedenti due test erano state cambiate le capacità degli effettori, in questo test mutano (ancora in modo mai sperimentato nel corso dell’evoluzione) i dati sensoriali. Anche in questo caso il sistema si è adattato adeguatamente.
Conclusioni
I risultati sembrano sostenere la tesi che condizioni simili portano a controlli motori simili. In meno di 4000 generazioni l’evoluzione simulata ha prodotto una famiglia di individui capaci di fissare e seguire un bersaglio mobile con movimenti coordinati occhi-testa simili a quelli dei sistemi biologici, e capaci di compensare mutamenti delle condizioni di funzionamento dei sensori e degli effettori.
Lo studio suggerisce inoltre che in questo tipo di simulazioni i risultati dipendono notevolmente dal livello di dettaglio nel simulare le caratteristiche ambientali e biologiche. Sicché, in questo tipo di simulazioni come nella robotica, l’attenzione dovrebbe essere spostata dal progettare i sistemi di controllo al progettare accuratamente il compito, l’ambiente, e le eventuali costrizioni fisiche e biologiche, lasciando al meccanismo evolutivo l’onere di individuare la soluzione.